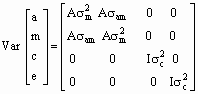|
Zootecnia Tropical 22(3):191-200. 2004 Parámetros genéticos para sobrevivencia al nacer en porcinos Rafael Galíndez1, Omar Verde2 y Gonzalo Martínez1
[1]
Universidad Central de Venezuela.
Facultad de Agronomía. Instituto de Producción Animal, Recibido y
aceptado dentro del contexto del XII Congreso de Producción e
Industria Animal 2004. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
RESUMEN Con la finalidad de estimar índices de herencia directos (h2d) y maternos (h2m), correlaciones genéticas entre efectos directos y maternos (rdm), heterosis y tendencias fenotípicas (Tf), genéticas (Tg) y ambientales (Ta), se analizaron 15308 registros de sobrevivencia de lechigadas al nacer, utilizando modelos lineales a través de dos metodologías: cuadrados mínimos (CM) y máxima verosimilitud restringida (REML). El modelo incluyó los efectos fijos: año y época de nacimiento, granja, número de parto, grupo racial de camada, las covariables: lechones nacidos vivos y peso de la camada al nacer, además de los efectos aleatorios de padre para CM y camada (directo) y madre (materno) para REML. Los análisis se realizaron por separado para razas blancas (RB = Large White, LW y Landrace, L) y razas de color (RC = Duroc, D y Hampshire, H). Los promedios no ajustados y ajustados resultaron en 94,53%, 93.54% para RB y 93.53%, 95.65% para RC. Los h2d utilizando CM y REML: 0.010 y 0.010; 0.004 y 0.010; 0.062 y 0.020; no estimable y 0.010 para LW, L, D y H, los h2m y rdm: 0.030 y 0.980; 0.020 y 0.950; 0.040 y 1.000, 0.070 y 1.000. La heterosis resultó estadísticamente no significativa (NS). Las Tg y Ta (P<0.01) corresponden a (0.580 y 0.684 en LW) y (-1.111 y 1.230 en L), las otras tendencias resultaron NS. Se concluye que las variaciones en la sobrevivencia de la población estudiada se deben en mayor proporción al ambiente y poco al efecto de los genes. Palabras clave: Índice de herencia, tendencias, correlaciones genéticas, cerdos, sobrevivencia al nacer. Genetics parameters of survival at birth in pigs SUMMARY Direct (h2d) and maternal (h2m) heritabilities, genetic correlation between direct and maternal effects (rdm), heterosis, phenotypic (Tf), genetic (Tg) and environmental (Ta) annual trends, were estimated using 15308 records of litter survival at birth (LSB), with two methodologies: Least Square (LS) and Restricted Maximum Likelihood (REML). The model included year and season of birth, farm, number of calving, litter breed group as fixed effects, number of piglets born alive and litter weight at birth as covariates. In addition, for LS sire, and for REML animal (direct) and dam (maternal) effects were included as random effects. The analyses were separated for white breeds (WB; Large White =LW and Landrace =L) and for color breeds (CB; Duroc = D and Hampshire = H). The adjusted and non-adjusted LSB averages were 94.53 and 93.54% for CB and 93.53 and 95.65% for WB. Estimates of h2d from LS and REML were 0.010, 0.010; 0.004, 0.010; 0.062, 0,020 and non-estimable, 0.010 for LW, L, D, and H, respectively. Estimates of h2m and rdm were 0.010, 0.98; 0.020, 0.95; 0.040, 1.000; and 0.07, 1.000 for LW, L, D, and H, respectively. There was not statistical evidence of heterosis for LSB. The Tg and Ta were 0.580 and 0.684 for LW, 1.11 and 1.230 for L, and were the only different from zero (P<0.01). The low estimates of heritabilities indicate that genetic effects are of little importance in comparison with the environmental effects for litter survival at birth in this swine population. Key words: Litter Survival at birth, heritability, genetic correlation, swine, annual trends. INTRODUCCIÓN La eficiencia económica de los sistemas de producción con cerdos depende, en gran manera, del producto que es llevado al mercado. Esto se traduce en la cantidad de individuos que son sacrificados para vender su carne. Por otra parte, toda unidad productiva debe producir sus reemplazos y, por ello, contar con más animales a la hora de seleccionar representa un aumento en la eficiencia de esta labor, puesto que la presión de selección que se ejerce será superior. Así pues, la ocurrencia de estos dos procesos estará supeditada a la sobrevivencia de los lechones tanto al nacer como al destete, estando esta tasa de sobrevivencia afectada por la acción del ambiente y de los genes que poseen los individuos. La alimentación en los períodos pre-parto de las marranas puede afectar de forma notoria el número de lechones que nacen vivos y, por consiguiente, los porcentajes de sobrevivencia de estas lechigadas. Asimismo, el diseño de las instalaciones de paritorio debe ser adecuado para evitar muertes durante y después del parto por aplastamiento. En las explotaciones porcinas es imprescindible cuidar de la sanidad del rebaño, ya que, las enfermedades representan la principal causa de muerte de los individuos cuando las condiciones sanitarias y los planes de vacunación no son los más adecuados. Sin embargo, puede existir una resistencia natural de los individuos a ciertas enfermedades como consecuencia de la acción de un grupo de genes. Por tal motivo, representa un reto a nivel de campo identificar los individuos que son superiores en tasa de sobrevivencia de sus hijos, para seleccionarlos y permitir que se reproduzcan y transmitan estos genes favorables a las siguientes generaciones y, por supuesto, descartar los individuos cuyo comportamiento productivo no sea satisfactorio al objetivo de producción. La presente investigación pretende estimar la proporción de la varianza observada que se debe a la acción de los genes; y de esta variación genética detectada, la proporción que corresponde a efectos genéticos aditivos directos y maternos; y los correspondientes efectos genéticos no aditivos expresados en la heterosis. MATERIALES Y MÉTODOS Se analizaron datos de sobrevivencia al nacimiento de dos granjas porcinas ubicadas en el estado Yaracuy. Los datos corresponden al período comprendido entre los años 1990 y 2000, ambos inclusive. Las condiciones de manejo en general eran similares para las dos granjas. Los animales consumieron un alimento balanceado producido por la misma empresa. En las granjas se llevó a cabo un plan de vacunación, así como un plan sistemático de desparasitaciones. Se aplica el manejo recomendado a los lechones (suministro de hierro, corte de cola y colmillos, etc.). En cuanto al manejo reproductivo se refiere, existió un estricto control de servicios, utilizando inseminación artificial en el 100% de los casos. Se definieron dos épocas de nacimiento: a) Época lluviosa: desde el 15 de Abril al 15 de Octubre y Época seca: desde el 16 de Octubre al 14 de Abril. La sobrevivencia al nacer de la camada se calculó dividiendo el número de lechones nacidos vivos entre el total de lechones nacidos, expresando el resultado en porcentaje. Se contó para los análisis con un total de 5793 registros para la raza Large White (LW), 4900 para Landrace (L), 2077 para los cruces LW con L, 1465 para Duroc (D), 217 para Hampshire (H) y 856 para los cruces D con H. Se realizaron análisis de varianza individuales para las cuatro razas y dos análisis combinados: uno para animales de razas blancas (Large White, Landrace y cruces recíprocos) y otro para animales de razas de color (Duroc, Hampshire y cruces recíprocos). Se calculó la heterosis mediante la diferencia entre el promedio de las camadas cruzadas y el promedio de las camadas de razas puras, expresándose los resultados en porcentaje. Los datos se analizaron por dos métodos: Cuadrados mínimos, en atención al número desigual de observaciones por clases, descrito por Harvey (1987). En análisis preliminares se incluyeron todas las interacciones, luego se descartaron para el análisis final aquellas interacciones que no resultaron significativas, quedando el modelo final de la forma siguiente: Modelo I: para estimar los promedios generales
donde: Yijklmno: porcentaje de sobrevivencia de la camada "o", nacida en el año "i", en la época "j", en una granja "k", proveniente de una madre que tiene "l" partos, de grupo racial de camada "m" , de un padre "n", que está presente dentro del grupo racial de camada "m". m = media teórica de la población. ai = efecto del año de nacimiento (i = 1990 a 2000). epj = efecto de la época de nacimiento (j = lluviosa, seca). grk = efecto de la granja (k = 1, 2). npl = efecto del número de parto de la madre (l = 1 a 8). grcm = efecto del grupo racial de camada (m = 1 a 4) (p/grc)nm = efecto del padre "n" dentro del grupo racial de camada "m". b1 = coeficiente de regresión lineal de la sobrevivencia sobre el número de lechones nacidos vivos (NLNV). b2 = coeficiente de regresión lineal de la sobrevivencia sobre el peso total de la camada al nacer (PCN). (a x np)il = interacción entre el año de nacimiento y el número de parto de la madre. єijklmno = residual con media cero y varianza s 2, normal e independientemente distribuido. Modelo II: para estimar el índice de herencia para sobrevivencia al nacer dentro de cada raza. Igual al modelo I, sin incluir el grupo racial de camada. Modelo III: para estimar las constantes fenotípicas para la sobrevivencia al nacimiento. Igual al modelo II, sin incluir el efecto de padre. En los análisis que involucran razas de color (Duroc y Hampshire) se excluyó el efecto de granja por estar presentes los grupos raciales en una. Los efectos de año y época de nacimiento, número de parto de la madre y granja son considerados fijos. Los efectos de padre y el residual se consideraron aleatorios. Máxima verosimilitud restringida (REML), utilizando MTDFREML (Boldman, et al. 1995): Para estimar la varianza genética aditiva directa, genética materna y la varianza del ambiente permanente se efectuaron análisis estadísticos utilizando dos modelos lineales aditivos mixtos univariados, con el objetivo de poder estimar los índices de herencia directo y materno, así como las correlaciones genéticas entre estos. En este sentido, los modelos utilizados fueron: Modelo IV: para estimar el índice de herencia y valores genéticos considerando el ambiente permanente.
donde: y = es el vector de observaciones de la variable respuesta. b = es el vector de efectos fijos: Año de Nacimiento (An): 1990 2000. Época de Nacimiento (En): Lluviosa - Seca. Número de Parto de la Madre (Np): 1 8. Granja (Gra): 1,2. Sólo para las razas Large White y Landrace. a = es el vector de efectos genéticos aditivos directos (aleatorio). m = es el vector de efectos genéticos maternos (aleatorio). c = es el vector de efectos ambientales permanentes (aleatorio). є = es el vector de efectos residuales. X = es la matriz de incidencia para los efectos fijos
Propiedades del Modelo: La esperanza de y es: E[y] = Xb La varianza del modelo es:
Donde A es la matriz de parentesco de todos los animales en la base de datos, I es una matriz de identidad de orden apropiado (número de madres para s 2c y número de registros para s 2e), s 2a es la varianza genética directa, s 2m es la varianza genética materna, s am es la covarianza entre los efectos genéticos directos y maternos, s 2c es la varianza de los efectos ambientales permanentes y s 2e es la varianza de efectos residuales. Modelo V: Igual al modelo IV, sin considerar el ambiente permanente. El criterio de convergencia utilizado fue cuando la varianza del 2Log de la verosimilitud fuese inferior a 10-6. Después de cada convergencia, el programa fue reiniciado en dos oportunidades, usando los estimados obtenidos anteriormente como valores iniciales, con el objeto de comprobar la convergencia. El efecto de la no inclusión del ambiente permanente en el análisis se cuantificó realizando una prueba de χ2 con un grado de libertad, para la diferencia entre los valores de los criterios de convergencia de los dos modelos. Se realizaron contrastes ortogonales para las comparaciones entre las lechigadas puras y cruzadas, con el fin de determinar la significancia estadística de la heterosis. RESULTADOS Y DISCUSIÓN Los promedios no ajustados y ajustados para la sobrevivencia al nacimiento corresponden a 94,53% y 93,54% y 93,53% y 95,65% para las razas blancas y de color, respectivamente. De manera general, estos valores de sobrevivencia son superiores a los señalados por Fahmy y Bernard (1971), Bereskin (1984), Gu et al. (1989), Quintero y Esparza (1992), Colmenares et al. (1994) y Chang (1997). Estas divergencias probablemente tengan su origen en las condiciones generales ambientales y de manejo entre los distintos trabajos citados. Por otra parte, la utilización de grupos raciales distintos en las investigaciones también puede introducir una fuente de variación en los resultados. Asimismo, la población analizada en este estudio estuvo sometida a programas estrictos de selección, por lo que el mayor valor promedio de sobrevivencia pudiera ser consecuencia de estos procedimientos. Otros autores (Fahmy et al., 1978, Peña y Verde, 1983, Ferrer, 1988, Southwood y Kennedy, 1990 y De Venanzi, 1998) encontraron valores similares (altos) para la sobrevivencia al nacimiento de camada, comportamiento que pudiera explicarse porque en la mayoría de estos trabajos, al igual que este, las poblaciones estuvieron sometidas a selección. Para la sobrevivencia al nacer, la mayoría de la variación fue causada por el ambiente. Ésta aseveración esta sustentada por los resultados mostrados en el Cuadro 1, donde todos los índices de herencia tanto directos como maternos son de baja magnitud (<0,10). Los valores de índice de herencia más altos fueron observados en los caracteres estudiados en la raza Duroc; empero, es necesario señalar que los resultados tienen altos errores estándar; el número de observaciones totales y el número de camadas por madre es muy bajo, lo que puede estar introduciendo imprevisiones en los cálculos. Por ejemplo, las correlaciones entre los valores directo y materno para las razas Duroc y Hampshire, son iguales a la unidad con errores de estimación superiores a uno. Otra evidencia de estos problemas de estimación se presenta en la raza Hampshire, en donde, por la metodología de cuadrados mínimos no es posible la estimación del índice de herencia directo. Los bajos índices de herencia señalados en este estudio concuerdan con los valores encontrados en la literatura (Rico y Gómez, 1982; López et al., 1988; Colmenares et al., 1994 y Van Arendonk et al., 1996). Por otra parte, en el último trabajo se reportan correlaciones bajas y negativas entre los efectos genéticos directo y materno; mientras que en el presente estudio las correlaciones son altas y positivas; lo que evidencia que la selección para este carácter incrementará su expresión tanto por la acción de los genes de efecto directo como por los genes de efecto materno. Sin embargo, esta ganancia entre generaciones se espera sea baja en vista de los valores bajos de los índices de herencia encontrados.
Se obtuvieron valores de heterosis de 1,14% y 0,34% (Cuadro 2) para los cruces que involucran razas blancas y de color, respectivamente. Estos estimados son muy bajos comparados con los que reporta Dieguez et al. (1996) de 3,5%, 3,5% y 3,0% para los cruces de las razas Yorkshire*Duroc, Yorkshire*Hampshire y Duroc*Hampshire, respectivamente. En el Cuadro 2 se puede observar que no existió efecto significativo del cruzamiento sobre el porcentaje de sobrevivencia al nacer. Esta afirmación es corroborada por la ausencia de diferencias estadísticas entre los promedios de los distintos grupos raciales. La acción ambiental pudiera estar enmascarando la expresión del vigor híbrido, ya que al someter los individuos a mejor manejo (mejores condiciones ambientales que se manifiestan en altos porcentajes de sobrevivencia), las diferencias en comportamiento entre grupos se reduce. No se debe dejar de lado que es probable que las diferencias genéticas en sobrevivencia entre las razas utilizadas en el estudio, sea menor de lo que hasta ahora se piensa.
Las tendencias genéticas y ambientales de la sobrevivencia al nacer para las razas Large White y Landrace, estimadas con las constantes obtenidas con el procedimiento de cuadrados mínimos resultaron significativas (P<0,01) (Gráficos 1 y 2). Los gráficos muestran el descenso ocurrido en el porcentaje de sobrevivencia a través de los años por causas genéticas, que en el caso de la raza Large White es de 0,5% en cada año y en la raza Landrace la reducción interanual en el porcentaje de sobrevivencia es de -1%. Las causas de este comportamiento evidentemente tienen que ver con la acción de los genes, producto de los planes de mejoramiento aplicados, que abarcan la selección y el cruzamiento. Es probable que la selección a favor del número de lechones nacidos vivos influya indirectamente en el tamaño de camada total (vivos y muertos), por lo que el incremento del número de lechones vivos traiga consigo aumento del número de lechones muertos y, por tal motivo, disminuye el porcentaje de sobrevivencia de estas lechigadas.
Las tendencias ambientales tienen sentidos opuestos a las tendencias genéticas. En las dos razas citadas, la acción del ambiente tendió a incrementar los porcentajes de sobrevivencia al nacimiento. Los cambios interanuales corresponden a 0,7% para la raza Large White y 1% para la raza Landrace. Los factores ambientales que afectan la sobrevivencia probablemente sean la alimentación, sanidad y las mejoras en el diseño de las instalaciones, sobre todo las instalaciones de maternidad. El antagonismo observado entre estas dos líneas de regresión ocasiona la anulación de ambos efectos, trayendo como resultado una tendencia fenotípica estadísticamente no significativa. Las otras regresiones estimadas con las constantes obtenidas por los métodos de cuadrados mínimos y máxima verosimilitud restringida que no han sido mencionadas, resultaron estadísticamente no significativas. CONCLUSIONES Los resultados evidencian un mayor efecto del ambiente sobre el carácter estudiado. Esta conclusión esta soportada en dos aspectos fundamentales: 1) La ausencia de efecto heterótico producto del esquema de cruzamiento aplicado en razas blancas y de color y 2) Los bajos índices de herencia, tanto directos como maternos estimados por las metodologías de cuadrados mínimos y máxima verosimilitud restringida. En tal sentido, se deben realizar mejoras sobre todo sanitarias y en las instalaciones para aumentar el porcentaje de sobrevivencia. Asimismo, el manejo aplicado a las marranas antes del parto debe ser favorable para no perjudicar a los lechones que están en formación en el vientre de éstas. En otro orden de ideas, es aconsejable realizar estudios de sobrevivencia en las cruzas entre cerdos de las razas blancas y de color, puesto que es de suponer que la semejanza genética entre estas es menor que la existente en las razas blancas o las de color entre sí, lo que pudiera conducir a una mayor expresión de la heterosis. BIBLIOGRAFÍA Bereskin, B. 1984. A genetic análisis of sow productivity traits. J. Anim. Sci., 59(5):1149-1163. Boldman, K., L. Kriese, L. Van Vleck, C. Van Tassell y S. Kachman. 1995. A manual for use of MTDFREML. A set of programs to obtain estimates of variance and covariances [Draft]. U. S. Department of Agricultue, Agriculture Research Service. pp 114. Chang, A. 1997. Comportamiento productivo predestete en una granja experimental porcina. Tesis Magister Scientarum. Facultades de Agronomía y Ciencias Veterinarias, Universidad Central de Venezuela.122 p. Colmenares, O., A. Chang y R. Álvarez. 1994. Factores genéticos y ambientales que afectan la mortalidad predestete en cerdos de diversas razas. VIII Congreso Venezolano de Zootecnia (San Juan de Los Morros, Venezuela). G018 (Resúmenes). De Venanzi, J. 1998. Caracterización de la producción en poblaciones porcinas puras y cruzadas. Tesis Magíster Scientarum. Facultades de Agronomía y Ciencias Veterinarias, Universidad Central de Venezuela. 224 p. Diéguez, F., T. Arias, G. Trujillo y I. Santana. 1996. Análisis de cruzamiento entre cerdos Yorkshire, Duroc y Hampshire en Cuba. Revista Computarizada de Producción Porcina 3(2). Disponible:http://www.sian.info.ve/porcinos/publicaciones /rccpn/Rev32/PACO. htm [Enero 2004]. Fahmy, M. y C. Bernard. 1971. Causes of mortality in Yorkshire pigs from birth to 20 weeks of age. Canadian J. Anim. Sci., 51(2):351-359. Fahmy, M., W. Holtmann, T. Macintyres y J. Moxley. 1978. Evaluation of piglet mortality in 28 two-breed crosses among eight breeds of pig. Animal Production, 26(3):277-285. Ferrer, R. 1988. Comportamiento de cerdas Yorkshire en un centro genético. XI Reunión ALPA (La Habana, Cuba). p.157 (Resúmenes). Gu, Y., Haley, C. y R. Thompson. 1989. Estimates of genetic and phenotypic parameters of litter traits from closed lines of pigs. Animal Production, 49(3):477-482. Harvey, W. 1987. Users guide for LSMLMW PC 1 version. Mixed model least squares and maximum likelihood computer program. 59 p. López, M., D. Guerra, J. Gómez y J. Dora. 1988. Parámetros genéticos de aspectos reproductivos del tamaño de la camada en cerdas. XI Reunión ALPA (La Habana, Cuba), pp.156 (Resúmenes). Peña, N. y O. Verde. 1983. Factores que influyen caracteres de sobrevivencia en lechones. IX Reunión ALPA (Santiago de Chile, Chile). GM-5 (Resúmenes). Quintero, M. y D. Esparza. 1992. Productividad predestete de lechones de razas Yorkshire, Landrace y sus cruces. VII Congreso Venezolano de Zootecnia GR-39 (Resúmenes). Rico, C. y J. Gómez. 1982. Factores genéticos y ambientales que influyen en el comportamiento reproductivo de la raza Duroc en Cuba. II Influencias en la mortalidad. Revista Cubana de Ciencia Agrícola, 16(1):35-41. Southwood, O. y B. Kennedy. 1990. Estimation of direct and maternal genetic variance for litter size in Canadian Yorkshire and Landrace swine using an animal model. J. Anim. Sci., 68(7):1841-1847. Van Arendonk, J., C. Van Rosmeulen, L. Janss y E. Knol. 1996. Estimation of direct and maternal genetic (co)variances for survival within litters of piglets. Livestock Production Science, 46(3):163 171. | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||